
创新性
1. 提出气候适应性害虫管理新策略:首次系统证明针对特定代谢酶的抑制剂可作为“防御诱导剂”,在预测的未来气候条件下(eCO2)发挥更佳效果,成功将气候变化的挑战(碳源增加)转化为植物防御强化的机遇。
2. 揭示代谢分支调控的精密性:不仅证实了“补偿性激活理论”,还精准定位了流向羟基苯甲酸衍生物这一高效抗虫分支的代谢流,为作物抗虫育种提供了新的代谢工程靶点。
3. 多维度验证机制:从田间相关的昆虫行为(EPG)到分子水平的互作(分子对接),研究提供了坚实且多层次的数据支持。
研究背景
工业革命以来,大气CO₂浓度从280 ppm升至420 ppm以上,高CO₂通过调节植物生理和害虫行为深刻影响植物-昆虫互作。eCO₂下,水稻光合作用增强但氮基防御物质(如生物碱)稀释,引发稻飞虱等刺吸式害虫的补偿性取食行为。苯丙烷代谢途径是植物次生代谢的核心网络,生成木质素、类黄酮及酚酸等防御化合物。eCO₂为植物提供丰富碳源,使得苯丙烷途径成为碳流关键通道,但其在抗虫防御中的调控机制尚不明确。本研究创新性地通过药理抑制剂(PA和MDCA)靶向干预C4H和4CL酶,探索其在eCO₂下能否通过重编程代谢流增强水稻抗虫性。
方法概述
实验设环境CO₂(aCO₂, 400 ppm)和高CO₂(eCO₂, 800 ppm)两组,在水稻幼苗(品种TN1)上施用300 μM PA或MDCA(以DMSO为对照)。通过电穿透图(EPG)记录稻飞虱取食行为,检测水稻ROS含量、PAL酶活性、基因表达及羟基苯甲酸衍生物含量,并分析稻飞虱生存率与解毒酶活性。分子对接模拟羟基苯甲酸衍生物与稻飞虱解毒酶(CarE、AChE、GST)的互作。数据采用双因素方差分析(Two-way ANOVA)和Mantel检验进行统计。
结果深度解析
1. 抑制剂显著抑制稻飞虱取食与生存,且eCO₂下效应倍增
EPG测试显示,PA和MDCA处理均缩短稻飞虱关键取食波形持续时间(N4a:韧皮部口针活动;N4b:韧皮部汁液吸食;N5:木质部口针活动)。在aCO₂下,PA使N4a、N4b和N5波形持续时间分别减少29.56%、40.46%和32.46%;而在eCO₂下,减少幅度高达85.34%、92.34%和88.44%。MDCA在eCO₂下亦显著抑制取食波形(N4a、N4b、N5分别减少62.76%、75.50%和66.18%)。生存率实验进一步证实,PA和MDCA处理显著降低稻飞虱生存率,且eCO₂下降幅最大(图1d)。这些结果说明eCO₂环境强化了抑制剂的抗虫效果。
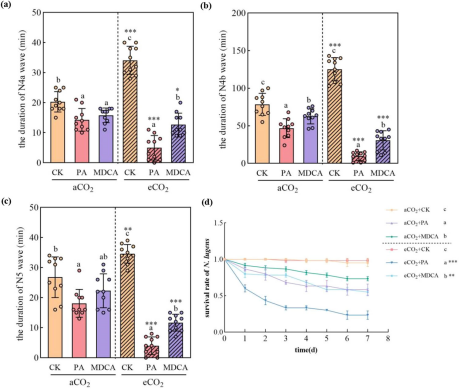
图(1)
图1 红色稻飞虱(BPH,Nilaparvata lugens)在施用 Piperonylic acid(PA)和 3,4-(甲氧基)肉桂酸(MDCA)处理的水稻植株上的取食行为及存活率,分别在常温CO₂(aCO₂)和升高水平CO₂(eCO₂)条件下。EPG测试记录了BPH在aCO₂和eCO₂条件下取食水稻植株时N4a(a)、N4b(b)和N5(c)波形的持续时间。在aCO₂和eCO₂条件下,N. lugens取食PA和MDCA处理水稻植株的存活率(d)。
2. eCO₂环境下,抑制剂诱发更强烈的ROS爆发
ROS是植物防御的早期信号分子。在未处理条件下,eCO₂水稻ROS含量低于aCO₂,可能与光合作用增强有关。但PA和MDCA处理逆转这一趋势:在eCO₂下,PA使ROS、H₂O₂和O₂⁻含量分别增加128.35%、149.62%和157.50%,MDCA亦显著提升ROS水平(102.51%)。表明抑制剂能高效利用eCO₂的碳源,触发氧化信号(图2)。
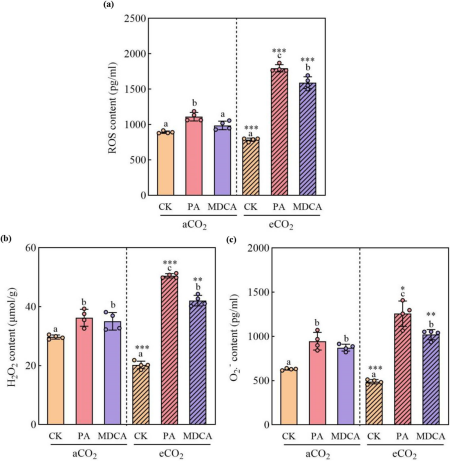
图(2)
图2 PA和MDCA处理对水稻在aCO₂和eCO₂条件下ROS含量的影响。PA和MDCA处理下水稻在aCO₂和eCO₂条件下的总ROS(a)、H₂O₂(b)和O₂⁻(c)含量。
3. ROS信号激活苯丙烷途径的核心引擎——PAL
PAL是苯丙烷途径的限速酶。PA和MDCA处理显著增强PAL酶活性,在eCO₂下PA使PAL活性提升106.32%。基因表达分析显示,PA在eCO₂下显著上调PAL1,3,4,6-9等基因表达(图3b,d,e,g-j),说明抑制剂通过ROS信号激活PAL介导的防御通路。
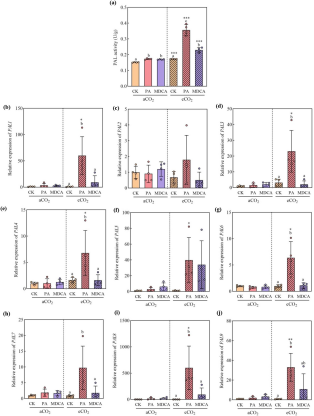
图(3)
图3 PA和MDCA处理对水稻在aCO₂和eCO₂条件下PAL活性(a)及PAL1–9基因相对表达量(b–j)的影响
4. 代谢流重定向:羟基苯甲酸衍生物大量积累
苯丙烷代谢途径存在分支竞争。抑制C4H或4CL(木质素合成关键酶)会阻断底物流向木质素分支,从而可能使其转向合成羟基苯甲酸衍生物。研究定量分析了6种羟基苯甲酸衍生物的含量。
图4的结果完美验证了这一假设。在eCO2下,PA和MDCA处理导致对羟基苯甲酸、焦儿茶酸、原儿茶酸、龙胆酸、没食子酸和香草酸等6种化合物的含量大幅上升。其中,PA处理对龙胆酸(增加573.82%)和香草酸(增加319.71%)的诱导尤为显著。曼特尔检验(Mantel test, 图4g)进一步证实,稻飞虱取食波形(N4b, N5)的持续时间与这些羟基苯甲酸衍生物的含量呈显著负相关,提示它们正是直接作用于昆虫的抗摄食物质。
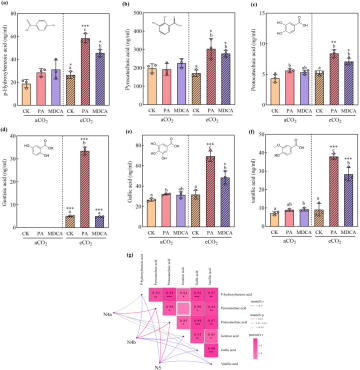
图(4)
图4 PA和MDCA处理对水稻植株在aCO₂和eCO₂下羟基苯甲酸衍生物含量的影响
5. 关键羟基苯甲酸衍生物具有强效抗摄食活性并影响生存
通过人工饲料选择实验,研究验证了上述化合物的直接功能。图5a-d显示,焦儿茶酸、龙胆酸、没食子酸和香草酸表现出极强的抗摄食活性,而对羟基苯甲酸和原儿茶酸效果较弱。图5g-j的生存实验表明,将这四种有效化合物分别以低(EC20)和高(EC80)浓度加入人工饲料后,能呈浓度依赖性地显著降低稻飞虱的生存率。
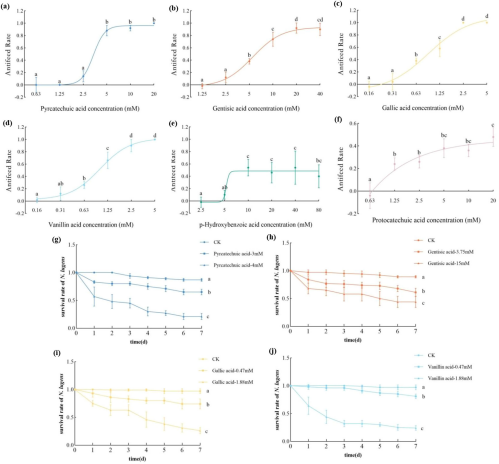
图(5)
图5 稻飞虱(Nilaparvata lugens)在饲喂含羟基苯甲酸衍生物的人工饲料时的取食偏好和存活率
6. 稻飞虱解毒酶应激反应揭示化合物毒性
昆虫的解毒酶(如乙酰胆碱酯酶AChE、羧酸酯酶CarE、谷胱甘肽S-转移酶GST、多功能氧化酶MFO)活性是其应对植物毒素的重要生理指标。研究发现,取食经PA/MDCA处理的水稻后,稻飞虱体内这四种解毒酶的活性均显著升高(图6a-d),且在eCO2下增幅更大。曼特尔检验(图6e)再次确认,解毒酶活性与四种关键羟基苯甲酸衍生物的含量显著正相关。通过人工饲料实验(图6f-h)进一步证实,正是这些衍生物直接诱导了AChE、CarE和GST的活性升高。这表明昆虫为代谢这些有毒物质付出了高昂的生理代价,最终导致死亡。
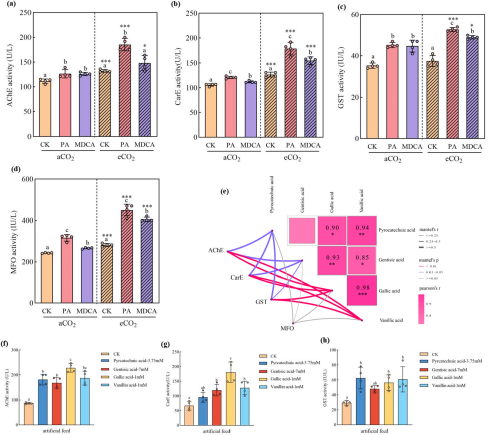
图(6)
图6 在aCO₂和eCO₂条件下,给予PA和MDCA处理的水稻植株及含有羟基肉桂酸衍生物的人工饲料对稻飞虱(Nilaparvata lugens)解毒酶活性的影响
7. 分子对接模拟揭示化合物与解毒酶的潜在直接互作
为从分子层面解释羟基苯甲酸衍生物如何影响解毒酶,研究进行了同源建模和分子对接。图7展示了四种衍生物与三种解毒酶(CarE, AChE, GST)在最优化结合模式下的结果。它们均能以较低的结合能(通常<-5 kcal/mol)与酶活性口袋内的多个氨基酸残基形成氢键等相互作用。例如,没食子酸与GST可形成4个氢键,结合能为-5.9 kcal/mol。这从理论上证明了这些化合物可以作为底物或抑制剂直接与稻飞虱的解毒酶结合,从而干扰其正常功能或诱导其过度表达。
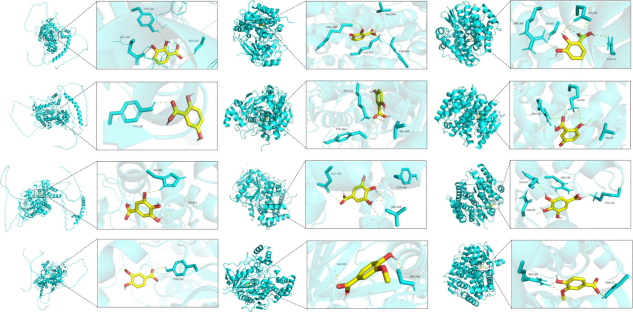
图(7)
图7 羟基苯甲酸衍生物与稻飞虱AChE、CarE和GST解毒酶的分子对接研究
结论
本研究揭示了“抑制剂→ROS爆发→PAL激活→羟基苯甲酸衍生物积累→昆虫取食抑制与死亡”的完整抗虫信号通路。在高二氧化碳浓度(eCO₂)环境下,充足的碳源进一步强化了苯丙烷代谢抑制剂PA和MDCA的防御诱导作用,其中PA因靶向上游酶C4H,在eCO₂条件下表现更优,可能与高效重定向碳流有关。羟基苯甲酸衍生物的积累是抗性增强的核心机制,而昆虫解毒酶活性升高则反映其生理应激并最终导致死亡。该研究表明,PA和MDCA通过激活苯丙烷代谢,可协同eCO₂下的碳源优势,显著增强水稻对稻飞虱的抗性,为开发气候智能型害虫管理策略提供了理论依据。未来需进一步评估抑制剂的田间持久性、环境安全性及其与生物防治的协同潜力。









